



Abstract
One of the important ways to identify features characteristic of different systems and organs of the human body is to study them using modern methods in vertebrates belonging to different phylogeny stages. The aim of the study was to investigate the spinal cord’s meninges of domestic ducks on histological and ultrastructural levels. The study was conducted on pieces of white matter of the spinal cord that were taken together with the meninges from domestic ducks. Araldite-Epon blocks were prepared from the samples according to the protocols accepted in electron microscopy, semi- and ultrathin sections were made on an ultratome and photographed under light and electron microscopes, respectively. Morphometric parameters of the structures were determined using a computer program for image analysis. The obtained data show that the dura mater of domestic ducks significantly differs from that of mammals due to detection of elastic fibers in their organization which length is measured in µm, as well as the transverse size 313,9-989,7 nm (626,0±16,71 nm). Between the elements of the fenestrated elastic membrane formed from elastic fibers are collagen fiber layers that are located in different directions. The presence of a boundary cell layer and a subdural space in the dura mater has been ultrastructurally refuted. According to the histotopography and the cellular and fibrillar structure of denticulate ligament of the spinal cord it has been determined that the ligament begins from the dura mater not from the pia matter of the spinal cord. The boundary between the cellular barrier layer and the reticular layer of the arachnoid matter is formed by a thin osmiophilic band which is created from the plasmalemma of leptomeningeal cells and the basal lamina located between them. The discovery of free elastic fibers in the arrangement of the dura mater and denticulate ligaments in domestic ducks together with the fenestrated elastic membrane may prevent pressure difference and abrupt changes in the position of the spinal cord during flight. With the use of modern technical facilities, it is possible transthecal injection of drugs or stem cells through denticulate ligament into the subarachnoidal space.
Full article
GİRİŞ
Sinir sisteminin müqayisəli anatomiyasının müasir metodların köməkliyi ilə tədqiqi beyin və onurğa beyninin strukturlarının funksiyaları, inkişaf və təkamül (adaptasiya) prosesləri haqqında yeni məlumatlar verməklə yanaşı, yalnız insan beyninə məxsus xüsusiyyətlərin aşkar olunmasında mühüm əhəmiyyətə malikdirlər [1, 2].
İstər mərkəzi, istərsə də periferik sinir sisteminin tərkibinə daxil olan demək olar ki, bütün strukturları birləşdirən ümumi cəhətlərdən biri onların quruluşca bir-birlərindən fərqlənən qişa elementləri ilə əhatə olunmalarıdır. Əldə olunan son məlumatlara görə beyin qişaları örtük funksiya ilə yanaşı, beyin-onurğa beyni mayesinin cərəyanında [3, 4], sinir elementlərinin histogenezi və onların normal fəaliyyəti üçün vacib olan interstisial mühitin tərkibinin sabit saxlanılmasında bilavasitə və sintez etdikləri bioloji aktiv maddələrin vasitəsilə iştirakları haqqında lazımi dərəcədə müasir məlumatlar alınmışdır [5, 6, 7]. İnsanın onurğa beynini örtən sərt və hörumçəktorunabənzər qişalarının elektron mikroskopik tədqiqi zamanı ilk dəfə olaraq hörumçəktorunabənzər qişanın xarici sədd və daxili tor qatından təşkil olunduğu qeyd edilmişdir [8]. Sədd qatları, aralarında kollagen liflərinin olmaması, bir-biri ilə sıx yerləşmiş, çoxlu miqdarda sıx əlaqələr vasitəsilə birləşmiş və tor qatdan fasiləli bazal səfhə ilə ayrılmaları ilə xarakterizə olunurlar.
Mövcud ədəbiyyat materiallarında (istər dünyanın inkişaf etdiyi ölkələrində dərc olunmuş dərslik və monoqrafiyalarda, istərsə də tədqiqat xarakterli məqalələrdə) baş beyin qişaları səviyyəsində S.Nabeshima və b. (1975) [9], onurğa beyni qişaları səviyyəsində isə F. Vandenabeele və b. (1996) [8] tərəflərindən müəyyən edilmiş sərt qişanın hüdudi hüceyrəvi qatı olduğu bir mənalı olaraq təkrar olunur [10,11]. İstinad olunan müəlliflər sərt qişanın hüdudi hüceyrəvi qatında yastılaşmış formaya malik, interdigitasiya üsulu ilə bir-birləri ilə birləşən, qatları arasında kollagensiz amorf maddə yerləşən, bazal səfhəsi olmayan, hüceyrəarası əlaqələrin az olması ilə xarakterizə olunan hüceyrə qatı yerləşdiyini qeyd edirlər. Qeyd etmək lazımdır ki, N. Wolf (2002) tədqiqat işində əldə etdiyi nəticələri və mövcud ədəbiyyat məlumatlarını ümumiləşdirərək, beyin qişalarının qatları, onların formalaşmasında iştirak edən hüceyrə tiplərinin tərkibi, hüceyrəarası əlaqələrin növləri, topoqrafik vəziyyətləri və s. xüsusiyyətlərinə görə ev quşları, məməlilər, o cümlədən insanlar arasında nəzərə çarpacaq fərq olmadığını qeyd etmişdir [12]. Ev qazlarında da sərt qişanın formalaşmasında iştirak edən strukturların ümumi quruluş planına və hüceyrə tiplərinə məməlilərlə müqayisədə nəzərə çarpacaq fərq aşkar edilməmişdir [13]. Onurğa beyninin sərt qişasına aid materiallar içərisində müzakirə olunası strukturlardan biri də onurğa beyninin onurğa kanalının stabilliyinin təmin olunmasında [14] və endoskopik cərrahi əməliyyatlar zamanı orientir rolunu oynayan mühüm əhəmiyyətə malik dişli bağlardır [15]. İnsanlarda dişli bağlar histoloji olaraq tədqiq olunsa da [16, 17, 18, 19], quşlarda dişli bağların formalaşmasında iştirak edən strukturlar ultrastruktur səviyyədə tədqiq olunmamışlar.
Göstərilənləri nəzərə alaraq, hazırki tədqiqat işinin məqsədi ev ordəklərində onurğa beyni qişalarının təşkilində iştirak edən hüceyrəvi və fibrilyar elementlərinin quruluş xüsusiyyətlərinin işıq və transmission elektron mikroskopik səviyyələrdə müəyyən edilməsidir.
TƏDQİQATIN MATERİAL VƏ METODLARI
Tədqiqat 6 baş ev ördəklərindən qişalarla birlikdə götürülmüş onurğa beyin ağ maddəsi tikələri üzərində aparılmışdır. Heyvanlar vivariumda standart şəraitdə saxlanılmışdır (hava temperaturu 22±2ºC, limitsiz su və yem istifadəsi). Bütün tədqiqatlar 18.03.1986 tarixli eksperimental və digər elmi məqsədlər üçün istifadə olunan Onurğalı Heyvanların Müdafiəsi Avropa Şurası Konvensiyasına uyğun olaraq aparılmışdır.
Götürülmüş tikələr 24 saat ərzində immersiya üsulu ilə fosfat buferində (pH=7,4) hazırlanmış 2%-li paraformaldehid, 2%-li qlütaraldehid və 0,1%-li pikrin turşusundan ibarət məhlulda fiksasiya edilmişdir. Nümunələr 1%-li osmium turşusu məhlulunda postfiksasiya edildikdən sonra artan dərəcəli etil spirti məhlullarında susuzlaşdırılaraq Araldit-Epon qətranlarında blok halına salınmışdır. Bloklardan EM UC7 (Leica, Almaniya) ultratomundan alınmış yarımnazik (1-2μm) kəsiklər metilen abısı, azur II və əsasi fuksinlə rənglənərək Primo Star (Zeiss, Almaniya) işıq mikroskopunda lazımi hissələrin şəkilləri EOS D650 (Canon, Yaponiya) rəqəmli fotokamera sistemi ilə çəkilmişdir. Eyni bloklardan alınmış 70-100 nm qalınlıqlı ultranazik kəsiklər əvvəlcə 2%-li uranil-asetat məhlulu, sonra NaOH-ın 0,1N qatılıqlı məhlulunda hazırlanmış 0,2%-li təmiz qurğuşun sitratla rənglənmişdir. Ultranazik kəsiklər 80-120 kv gərginlik altında JEM-1400 (Joel, Yaponiya) transmission elektron mikroskopunda tədqiq olunaraq elektronoqramlar çəkilmişdir.
Tədqiq olunan strukturların morfometrik parametrləri Almaniyanın «Olympus Soft İmaging Solutions Gmbh» şirkəti tərəfindən hazırlanmış təsvirin analizi kompyuter proqramı (The TEM imaging platform) vasitəsilə TİF formatında çəkilmiş elektronoqramlarda müəyyən edilmişdir.
NƏTİCƏLƏR
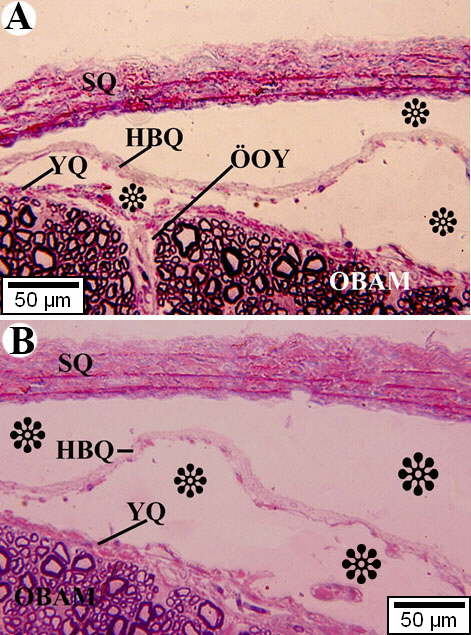
Ev ördəklərinin onurğa beyninin qişaları və ağ maddələrinin histotopoqrafiyası şəkil 1A və 1B-də nümayiş etdirilmişdir. Göründüyü kimi sərt qişa (SQ) ilə yumşaq qişa (YQ) arasında zolaq şəklində yerləşən hörümçəktorunabənzər qişanın ətrafinda (HBQ) rənglənməmiş hissələrin (ulduzlarla işarə olunublar) olması, ilk baxışdan onların subdural (yuxarıda) və subaraxnoidal (aşağıda) boşluqlara aid olduqları kimi qəbul oluna bilər. Şəkil 1A-da nəzər diqqəti cəlb edən yumşaq qişa elementlərinin onlarla birlikdə damarların onurğa beyninin ön orta yarığına (ÖOY) daxil olmasıdır. Onurğa beynin ağ maddəsinin (OBAM) təşkilində müxtəlif diametrli mielinli sinir liflərinin təşkil olunduğu aydın görünür.
Subdural boşluq kimi (SDB) təsvir olunan rənglənməmiş sahələrin həqiqətdə hörümçəktorunabənzər qişanın fraqmentasiyası nəticəsində meydana çıxdığı ultrastruktur olaraq aşkar olunmuşdur. Şəkil 2A-da nümayiş etdirilən elektronoqrammada göründüyü kimi hörümçəktorunabənzər qişanın ox başı ilə göstərilən fraqmentinin sərt qişanın daxili qatında yerləşən kollagen lifi dəstələri səviyyəsində qaldığı aydın görünür. Şəkil 2B-də isə göründüyü kimi yuxarıda təsvir olunan fraqmentin sola doğru davamının hörümçəktorunabənzər qişanın tərkibində (ox ucları ilə göstərilib) qaldığı aydın görünür. Beləliklə, subaraxnoidal boşluq kimi (SAB?) təsvir olunan rənglənməmiş sahənin hörümçəktorunabənzər qişanı elektron mikroskopik tədqiqat üçün işlənməsi zamanı meydana çıxan artefakt kimi qəbul etməyə tam əsas verir.
Onurğa beynin sərt qişasının hüceyrəvi və fibrilyar strukturlarının histotopoqrafiyası və ultrastruktur quruluş xüsusiyyətləri şəkil 3-də nümayiş etdirilmişdir. Ulduzla işarə olunmuş artefakt subdural boşluğların sərhədlərində genişlənmiş dənəli endoplazmatik şəbəkə sisternaları yerləşən fibroblastların periferik (şəkil 3A) və mərkəzi hissələrinin (şəkil 3B) yerləşdikləri nümayiş etdirilmişdir. Bu elektronoqrammalarda nəzər diqqəti cəlb edən müxtəlif ölçüyə, formaya və gedişə malik elastiki liflərdən təşkil olunmuş pəncərəli elastik zarın (PEZ işarə olunub) yerləşmələridir. Elektronoqrammalardan göründüyü kimi pəncərəli elastiki zarın liflərin arasında dalğavari gedişə malik kollagen lifi səfhələri (KLS) yerləşirlər. Şəkil 3C-də sərt qişa daxilində fibroblast hüceyrəsinin nüvəsi (Fbn) ilə bərabər nüvəciyi yerləşir. Bu elektronoqrammada kollagen lifi səfhələri daxilində müxtəlif istiqamətlərdə gedən kollagen liflərin yerləşməsi nəzər diqqəti cəlb edir. Şəkil 3C-də artefakt subdural boşluq (ulduzla işarə olunub) sərt qişanın digər hissələrində də müəyyən edilən fibrositin nüvə (FN) yerləşən mərkəzi və periferik hissələri əhatə olunmuşdur. Fibrositin böyüdülmüş fraqmenti elektronoqrammanın yuxarı sol bucağında yerləşmişdir. Göründüyü kimi fibrositin nüvəsinin periferik hissəsində nisbətən nazik heteroxromatin toplantıları arasında nüvə dəlikləri yerləşir. Ümumiyyətlə, nukleoplazmada üstünlük təşkil edən euxromatin törəmələridir. Fibrositin nüvə ətrafı hissəsində ikincili lizosomlar, dənəli endoplazmatik şəbəkənin elementləri və ayrı-ayrı ribosomlar yerləşir. Qeyd etmək lazımdır ki, əksərən sərt qişa ilə hörümçəktoruna bənzər qişanın sədd qatı arasında kollagen lifləri aşkar olunurlar. Şəkil 3Ç-də ulduzla işarə olunmuş boşluqlar sərt qişa elementlərindən əsasən kollagen lifi dəstələri vasitəsilə ayrılır. Belə ki, fibrositin nüvəsi yerləşən mərkəzi hissəsi (FN) və onun periferik hissələri hər iki tərəfdən kollagen lifləri ilə əhatə olunmuşdur. Elektronoqramma boyunca fibrositlərin nüvə və periferik hissələrin müxtəlif səviyyədə yerləşdiyini görürük. Digər xarakterik xüsusiyyət pəncərəli elastiki zarın təşkilində iştirak edən böyük ölçüyə malik elastiki liflərin sərt qişanın daxili qatında yerləşməsi aşkar olunmuşdur.
Onurğa beynin dişli bağının beynin sərt qişasından başlandığı yeri sonuncunun gedişinin işıq mikroskopik və dişli bağın tərkibində iştirak edən elementlərin elektron mikroskopik görüntüsü şəkil 4-də nümayiş etdirilmişdir. Şəkil 4A-da göründüyü kimi metilin abısı ilə rənglənən yarım nazik kəsiyin təxminən mərkəzi hissəsindən başlayan dişli bağ sola aşağıya doğru gedərək onurğa beynin yumşaq qişa (YQ) elementlərinə çatır. Aparılan tədqiqatlar bir mənalı göstərir ki, subaraxnoidal bucaq nahiyəsindən başlayaraq dişli bağ elementləri ancaq hörümçəktorunabənzər qişanın torlu qatının təşkilində iştirak edən leptomeningeal hüceyrələrlə hər tərəfdən əhatə olunurlar. Bunun nəticəsidir ki, dişli bağların təşkilində iştirak edən lifli və hüceyrəvi strukturlar sol (şəkil 4B) və sağ (şəkil 4C) tərəflərində yerləşən bir və ya iki qat leptomeningeal hücerələr (LMH) vastəsilə SAB cərəyan edən beyin-onurğa beni mayesindən ayrılırlar.
Dişli bağın başlanğıc hissəsində (şəkil 5A) sərt qişada olduğu kimi (bax şəkil l və səkil 2) pəncərəli elastiki zarın (PZF) təşkilində iştirak edən elastiki liflərin aralarında kollagen lifi səfhələri (KLS) yerləşirlər. Təsvir olunan elektronoqrammada digər diqqəti cəlb edən onun diaqonalı boyu yerləşən elastiki lifin mərkəzi hissəsinin iki şaxəyə bölünməsidir. Dişli bağların gedişi boyu isə elastiki liflərin köndələn kəsikləri aralarında fibrisitlərin mərkəzi və periferik hissələri ilə əhatə olunmuş kollagen lifi dəstələri (KLD) yerləşirlər (şəkil 4B -4C və şəkil 5B). Şəkil 5C-də maraqlısı odur ki, dişli bağın təşkilində iştirak edən şüalı quruluşa malik olan fibrosit sitoplazmasından başlayan çıxıntıların kollagen lifi dəstələrini əhatə etməklə yanaşı elastiki liflərlə də sıx əlaqədə olurlar. İlk baxışda elastiki liflərin kollagen lifləri arasında yerləşdiyi təsəvvürü yaranır.
Ancaq təsvir olunan elektronoqrammanın miqyasını artıran zaman elastiki liflərin fibrositin mərkəzi hissəsinin (FN) ətrafında, tam əksəriyyətinın isə fibrositlərin çıxıntıları ilə sıx təmas şəklində yerləşdikləri müəyyən edilir.
Qeyd etmək lazımdır ki, istər sərt qişaların özlərinin, istərsə də dişli bağların tərkibində sərbəst şəkildə və ya damarların ətrafında perisitlərlə (P) yanaşı mononuklear hüceyrələr aşkar olunurlar (şəkil 5Ç). Göründüyü kimi elektronoqrammanın sol aşağı hissəsində yerləşən mononuklear hüceyrə (MNH) nüvəsində endotel və perisit hüceyrələri ilə müqayisədə heteroxromatinin miqdarının çox olması, nüvə dəliyi kompleksinin aydın görünməsi, sitoplazmasında sentriolun, müxtəlif sıxlıqlı ribosomların və tək-tək pinositoz qovuqcuqların olması ilə fərqlənir.
Hörümçəktoruna bənzər qişanın (HTQ) təşkilində iştirak edən leptomeningeal hüceyrənin və kollagen lifi dəstələrinin histotopoqrafiyası və ultrastruktur quruluş xüsusiyyələri şəkil 6-da nümayiş etdirilmişdir. Şəkil 6A-da hörümçəktorunabənzər qişanın həm hüceyrəvi sədd (HQHSQ), həm də tor (HQTQ) qatlarının təşkilində leptomeningeal hüceyrələrin çıxıntıları ilə (LMHÇ) yanaşı, nüvə yerləşən (LMHN) mərkəzi hissələri aydın görünürlər. Leptomeningeal hüceyrələrin nüvə örtüyü ilə əlaqəli hissələrində nazik ölçüyə malik heteroxromatin strukturları yerləşdiyi halda, nukleoplazmanın əsas kütləsində euxromatin strukturları və bəzən nüvəciklər yerləşirlər. HQHSQ başlanğıc hissəsində və HQTQ qatında yerləşən leptomeningeal hüceyrələrin sitoplazmasında örtüklü, örtüksüz pinositoz qovuqcuqlar, dənəli endoplazmatik şəbəkə sisternaları, Holci apparatı, tək tək lizosomlar və mitoxondrilər aşkar olunurlar. Ələlxüsus torlu qatın təşkilində iştirak edən hüceyrələrin sitoplazmalarında sadalanan əlamətlərın olması, onların sintetik aktivliyinin yüksək olduğunu göstərir. Qeyd etmək lazımdır ki, HQHSQ-ı ilə HQTQ-i sərhəddində qonşu hüceyrələrin plazmolemmaları ilə onların arasında yerləşən bazal səfhə elementlərinin sıx yerləşmələri nəticəsində tünd osmiofil (şəkil 6A və B-də enli oxlarla işarələniblər) struktur formalaşır. Təqdim olunan elektronoqrammada nəzər diqqəti cəlb edən hörümçəktorunabənzər qişanın hüceyrəvi sədd və torlu qatlarının təşkilində iştirak edən leptomeningeal hüceyrələrin bir-birilərinə söykənən hissələrində incə quruluşa malik desmosomların, kommunikasion və sıx əlaqələrin müəyyən edilməsidir.
Şəkil 6 B-də nümayiş etdirilən HBQ fraqmentinin fərqli xüsusiyyəti onun ancaq leptomeningeal hüceyrələrin periferik hissələrindən təşkil olunmasıdır. Göründüyü kimi HQTQ-nın təşkilində iştirak edən leptomeningeal hamısının köndələn ölçülərinin 100 nm-dən az olmasına baxmayaraq, sitoplazmalarında dənəli endoplazmatik şəbəkələrin, ribosomların və sitoskelet elementlərinin yerləşdikləri müəyyən edilir. Maraqlısı odur ki, HQHSQ-nın ilk qatlarında müxtəlif orqanellərin və sıx yerləşən sitoskelet strukturlarının aşkar olunmasına baxmayaraq səthi qatda yerləşən leptomeningeal hüceyrələrin çıxıntılarının köndələn ölçülərinin azalması ilə yanaşı, sitoplazmalarında orqanellər demək olar ki aşkar olunmur (ox başları ilə işarə olunublar). Bu əlamət digər çoxqatlı epitel örtüklərində olduğu kimi səthi qatda yerləşən hüceyrələrin fuksional aktivliyin azalması kimi qiymətləndirmək olar.
Onurğa beynin ağ maddəsi və onun əhatəsində yerləşən yumşaq qişa elementlərinin və hissəvi olaraq HQTQ aid olan strukturların elektron mikroskopik görüntüsü şəkil 7-də nümayiş etdirilib. Qeyd etmək lazımdır ki, hörümçəktorunabənzər qişa ilə yumşaq qişa arasında dəqiq sərhəd təyin etmək mümkün olmur. Yeganə onu qeyd etmək olar ki, onurğa beyni ağ maddəsini xaricdən əhatə edən xarici (hüdudi) qliya zarının təşkilində iştirak edən lifli astrositlərin bazal səfhələrinə (şəkil 7A və C-yə qədər ox başları ilə işarə olunub) söykənən strukturlar yumşaq qişaya aid edilir. Şəkil 7B-də yumşaq qişanın təşkilində iştirak edən leptomeningeal hüceyrələrin çıxıntısında Holci aparatı sisternalarının toplantıları, dənəli endoplazmatik şəbəkənin sisternaları aşkar olunmuşdur. Digər nəzər diqqəti cəlb edən leptomeningeal hüceyrələrin ətrafında və onların çıxıntıları arasında kollagen lifləri ilə yanaşı nisbətən nazik ölçüyə malik elastiki liflər yerləşmişdir. Xarici hüdudi qliya zarının və subpial boşluğun (SPB) ətrafında yerləşən yumşaq qişanın təşkilində iştirak edən leptomeningeal hüceyrələrin periferik çıxıntıları arasında sıx əlaqələrə məxsus bitişmə nöqtələrinin və kommunikasion əlaqələrə xas quruluş aşkar olunmuşdur (şəkil 7C). Şəkil 7Ç-də tərkibində ara filamentlər (AF-la işarə olunub) aşkar olunan üçqat lifli astrosit ayaqcıqlarından formalaşan XHQZ-n ətrafında yumşaq qişanın sıx yerləşən leptomeningeal hüceyrələrinin çıxıntıları və kollagen lifli dəstələrindən təşkil olunması görsənir.
MÜZAKİRƏ
Ədəbiyyat icmalında qeyd olunduğu kimi beyin qişaları səviyyəsində bioloji mayelərin cərəyanında iştirak edə bilən sərbəst subaraxnoidal boşluğun olduğu göstərilir. Bizim kafedrada aparılan histoloji və ultrastruktur tədqiqatlar məməlilərdə (o cümlədən insanda) baş beyin qişalarında sərbəst subaroxnoidal boşluğun mövcud olmadığı həmişə qeyd edilmişdir. Hal-hazırki tədqiqat işində ev ördəklərinin baş beyin qişaları səviyyəsində istər histoloji, istərsə də ultrastruktur səviyyədə rənglənməmiş sahələrin olduğu bir daha öz təsdiqini tapmışdır.
Qeyd olunanlara baxmayaraq, hələ də beyin qişalarının təşkilində iştirak edən strukturların qarşılıqlı əlaqələri, ultrastruktur quruluş xüsusiyyətləri və müəyyən funksiyaların yerinə yetirilməsindəki rolları haqqında fikir ayrılıqları qalmaqda davam edir. Baş və onurğa beyninin sərt qişası tərkibində müxtəlif funksional aktivliyə malik fibrositlər və mononuklear hüceyrələrlə yanaşı, ultrastruktur olaraq nüvə-sitoplazma münasibətləri yüksək olan, periferik lokalizasiyaya malik heteroxromatin strukturlarının üstünlük təşkil etməsi və sitoplazmasında orqanellərin sayının kəskin azlığı ilə fərqlənən hüceyrələr də müəyyən edilmişdir. Müvafiq ultrastrukturlara malik hüceyrələrin immunhistokimyəvi olaraq nestin, Sox2, doublecortin, vimentin aktivliyi olan, in vitro və transplantasiyadan sonra in vivo sinir hüceyrələri istiqamətində differensasiya etmə imkanlarına malik sələf hüceyrələri olduqları müəyyən edilmişdir [20, 21].
Bununla birlikdə, istər tədqiq olunan məməlilərdə, istərsə də quşlarda baş beyin və onurğa beyninin sərt qişaları ilə hörümçəktorunabənzər qişa elementləri arasında fasiləsiz hüceyrə qatının olması bizim tərəfimizdən təsdiq olunmamışdır.
Tədqiqat obyektlərinin hamısında histoloji, immunhistokimyəvi və elektron mikrospik olaraq sərt və hörümçəktorunabənzər qişa elementləri arasında bioloji mayelərin cərəyanında iştirak edə biləcək subdural boşluğun mövcudluğu təkzib edilmişdir. Qeyd etmək lazımdır ki, bəzi alimlər tərəfindən epidural boşluqlar kimi subdural boşluqların da ancaq patoloji proseslər zamanı meydana çıxması fikirləri ilə tam razılaşmaq olar [22, 23]. Nəzarət qrupunda sərt qişa ilə hörümçəktorunabənzər qişa elementləri arasında struktur elementləri aşkar olunmayan rənglənməmiş sahələrin qeyd edilməsi preparatların parafin və araldit-epon bloklarının hazılanması zamanı subaraxnoidal boşluqda cərəyan edən beyin-onurğa beyni mayesinin dehidritasiyası nəticəsində meydana çıxan artefakdır.
Tədqiq olunmuş məməlilərlə müqayisədə [8] ev quşlarında onurğa beynini əhatə edən sərt qişanın fibrilyar quruluşa malik strukturların tərkibləri və morfometrik göstəriciləri nəzərə çarpacaq dərəcədə fərqlənirlər. Belə ki, istər ev ördəklərinin, istərsə də ev qazlarının onurğa beyinlərini əhatə edən sərt qişanın təşkilində boylama ölçüləri mkm-lərlə ölçülən, diametrləri orta hesabla 313,9-989,7 nm (626,0±16,71 nm) olan elastiki liflər yerləşirlər. Təsvir olunan sərt qişaların köndələn kəsiklərində müxtəlif ölçülü elastiki liflər qatı kollagen liflərindən təşkil olunmuş qatlarla əhatə olunaraq, aortanın orta qişasının təşkilində iştrak edən, pəncərəli elastiki zarlara bənzər struktur formalaşdırırlar. Ancaq quşların onurğa beyninin sərt qişası tərkibində pəncərəli elastiki zarın formalaşması, onların potensial olaraq quşların uçmaları zamanı onurğa kanalında onurğa beyninə aid strukturların topoqrafik vəziyyətlərinin təsbit olunmasında (sabit saxlanılmasında) mühüm əhəmiyyətə malik olmalarını söyləməyə əsas verir.
Əldə olunmuş məlumatlar bir mənalı göstərir ki, sərt qişanın hüdudi hüceyrəvi qatına aid edilən hüceyrələrin hörümçəktorunabənzər qişanın sədd qatının leptomeningeal hüceyrələri və bir-birləri ilə sıx təmasda olmaları, kommunikasion, desmosomal, ələlxüsus, araxnoidal səddin morfoloji göstəricisi olan sıx əlaqələrlə [24] birləşməsi onların da sədd qatının tərkib hissəsi olduğunu göstərir. İlk qatından başlayaraq sitoskelet elementlərinin kəskin sıxlaşması, nüvənin təşkilində iştirak edən xromatin strukturlarının kondensasiyası, səthi qat hüceyrələrində nüvənin ümumiyyətlə aşkar olunmaması, orqanellərdən cəmi tək-tək ribosomların və daraqların destruksiya vəziyyətində olan mitoxondrilərin olması epitelial quruluşa malik gedən buynuzlaşma prosesinin əlaməti kimi qəbul edilməlidir. Təsvir olunan quruluşa malik hüceyrələr ağ siçovulların və ev quşlarının sərt qişaları tərkibində aşkar edilmirlər.
Mövcud ədəbiyyat materiallarında daimi olaraq müzakirə olunan məsələlər içərisində mərkəzi və periferik sinir sistemlərinə aid qişa elementlərinin müvafiq olaraq beyin-onurğa beyni və endonevral mayelərinin cərəyanında iştirakları həmişə, elə indi də tədqiqatçıların diqqət mərkəzindədir. Məsələnin tarixinə varmadan qeyd etməliyik ki, beyin-onurğa beyni mayesinin cərəyanına həsr olunmuş sonuncu elmi tədqiqat işlərindən birinə müraciət etməyi lazım bilmişik. Bir qrup İsveçrəli alimlər [25] xəlbir sümüyünün üfiqi (xəlbir) səfhəsi səviyyəsində qoxu siniri lifi dəstələrinin, beyin qişalarının və limfa damarlarının qarşılıqlı əlaqələrini tətqiq etmişlər. Məlliflər xəlbir səfhə dəlikləri səviyyəsində hörümçəktorunabənzər qişanın sədd qatında fasilələrin olduğunu aşkar etmişlər. Bu isə qoxu siniri dəstələrinin xəlbir sümüyü dəliyindən keçən zaman sədd rolunu oynayan qişa elementləri ilə örtülmürlər. Nəticədə subaraxnoidal boşluqda yerləşən beyin-onurğa beyni mayesi qoxu siniri dəstələri ilə birlikdə burun boşluğunun qoxu nahiyəsinin selikalti qişasının xüsusis səfhəsində yerləşən limfa kapilyarlarına daxil olular.
Əldə olunan məlumatlar bir mənalı göstərir ki, tamamilə oxşar vəziyyət onurğa beyninin dişli bağları səviyyədə müşahidə olunur. Belə ki, dişli bağların onurğa beynin sərt qişasından başlandığı səviyyələrdə hörümçəktorunabənzər qişanın sədd qatına aid strukturları aşkar olunmur. Dişli bağlar subaraxnoidal boşluqda ancaq hörümçəktorunabənzər qişanın tor qatına aid bir və ya iki qat leptomeningeal hüceyrələrin (bioloji sədd funksiyssına malik olmayan) əhatəsində onurğa beyninin yumşaq qişa səviyyəsinə çatırlar (bax yuxarı). Bununla da, subaraxnoidal boşluğun özündə və leptomeningeal hüceyrə qatları arasında yerləşən beyin-onurğa beyni mayesinin dişli bağların kollagen lifləri boyunca nəql olunub sərt qişa daxilində yerləşən limfa damarlarına daxil olmalarına heç bir maneə qalmır [26].
S. Proulxun (2021) dərc etdirdiyi icmal xarakterli məqalədə beyin-onurğa beyni mayesinin cərəyanında görmə və onurğa beyni sinirlərini əhətə edən qişa elementlərinin də iştirak etdikləri nümayiş olunmuşdur [27]. Qeyd etmək lazımdır ki, bizim kafedra əməkdaşları tərəfindən [28] ağ siçovulların görmə sinirlərinin subaraxnoidal bücağı səviyyəsində hörümçəktorunabənzər qişanın atmaları arasında, ölçüləri 8-35 mkm olan likvordaşıyıcı kanalların (termin M. Baron və M. Mayorova tərəfindən 1982-ci ildə irəli sürülmüşdür) olduğu nümayiş etdirilmişdir [29]. Qeyd etmək lazımdır ki, ardıcıl yarımnazik kəsiklərin müqayisəli tədqiqi zamanı təsvir olunan likvordaşıyıcı kanalların mənfəzlərinın göz yuvasının birləşdirici toxuma elementləri ilə birbaşa əlaqədə olduqları dəqiqləşdirilmişdir.
Hal-hazırki tədqiqat işində onurğa beyni sinirlərinin ön köklərinin spinal qanqliona söykəndiyi yerdə formalaşan subaraxnoidal bucaqlar səviyyələrində hörümçəktoruna bənzər qişalardan başlayan atmalarla əhatə olunmuş likvordaşıyıcı kanalar aşkar olunmuşdur. Yekun olaraq qeyd etmək lazımdır ki, beyin-onurğa beyni mayesinin subaraxniodal boşluqdan dişli bağlar və subaraxnoidal bucaq nahiyələrində yerləşən likvordaşıyıcı kanallar vasitəsilə beyin-onurğa beyni mayesinin onurğa beyninin sərt qişası tərkibində olan venoz və limfa damalarına daxil olmalarının morfoloji əsasları mövcudur.
Son zamanlar dərc olunan elmi tədqiqatların əksəriyyətində hörümcəktorunabənzər qişa səviyyəsində formalaşan bioloji səddi qan-araxnoidal sədd (blood-arachnoid barrier) adlandırırlar [30, 31, 32, 33]. Qeyd etmək lazımdır ki, hörümçəktorunabənzər qişa səviyyəsində damar toru olmadığından onun hüceyrəvi sədd qatı qanla yox, sərt qişa damarlarından xaric olan intestisial maye ilə təmasda olurlar. Beləliklə, hörümçəktorunabənzər qişanın hüceyrəvi sədd qatının sərt qişanın intestisial və subaraxnoidal boşluqda cərəyan edən beyin-onurğa beyni mayelərinin aralarında yerləşdiyini nəzərə alaraq, formalaşan bioloji səddi sədəcə olaraq araxnoidal sədd adlandırılması daha məqsədəuyğundur [34, 35, 36].
Təsvir olunanlar, alimlər tərəfindən təklif olunan araxnoidal səddin təşkilində iştirak edən leptomeningeal hüceyrələr arasında formalaşan sıx əlaqə komplekslərinin balıq və siçovullarla müqayisədə quşlarda daha mürəkkəb quruluşa malik olması fikirləri ilə razılaşmağa əsas verir [24].
YEKUN
Əldə olunan məlumatlar göstərir ki, ev ördəklərində sərt qişanın məməlilərdən kəskin fərqi onlarda elastiki liflərin aşkar olunmasıdır. Sərt qişada elastiki liflərin iştirakı ilə formalaşan pəncərəli elastiki zar elementləri arasında müxtəlif istiqamətdə yerləşən kollagen lifi səfhələri, dişli bağlarda isə kollagen lifi dəstələri yerləşir. Sərt qişanı hörumçəktorunabənzər qişadan ayıran hüdudi hüceyrə qatının və subdural boşluğun mövcudluğu ultrastruktur olaraq təkzib edilmişdir.
Onurğa beyni qişalarının dişli bağlarının histotopoqrafiyasına, hüceyrəvi və fibrilyar elementlərin tərkibinə görə onurğa beyninin yumşaq qişasından yox, sərt qişadan başladığı müəyyən edilmişdir. Hörumçəktorunabənzər qişanın hüceyrəvi sədd qatı ilə tor qatı arasında sərhəd leptomeningeal hüceyrələrinin plazmalemmaları və onların arasında yerləşən bazal səfhənin bir-birinə sıx yerləşmələri nəticəsində formalaşan nazik osmiofil zolaq təşkil edir.
Figures

Keywords
References
1. Takemura H., Pestilli F., Weiner K.S. Comparative neuroanatomy: Integrating classic and modern methods to understand association fibers connecting dorsal and ventral visual cortex // Neurosci Res., 2019, 146: 1-12, https://doi.org/10.1016/j.neures.2018.10.011
2. Toossi A., Bergin B., Marefatallah M., Parhizi B., Tyreman N., Everaert D.G., Rezaei S., Seres P., Gatenby J.C., Perlmutter S.I. Comparative neuroanatomy of the lumbosacral spinal cord of the rat, cat, pig, monkey, and human // Sci Rep., 2021, 11(1): 1-15, https://doi.org/10.1038/s41598-021-81371-9
3. Siegenthaler J.A., Pleasure S.J. We have got you 'covered': how the meninges control brain development // Current Opinion in Genetics and Development, 2011, 21(3): 249-255, https://doi.org/10.1016/j.gde.2010.12.005
4. Weller R., Sharp M., Christodoulides M., Carare R.O., Mollgard K. The meninges as barriers and facilitators for the movement of fluid, cells and pathogens related to the rodent and human CNS // Acta Neuropathologica, 2018, 135(3): 363-385, https://doi.org/10.1007/s00401-018-1809-z
5. Liddelow S., Dziegielewska K., Ek J., Mark D. Habgood, Bauer H., Bauer H.-C., Lindsay H., Wakefield M.J., Strazielle N., Kratzer I. Mechanisms that determine the internal environment of the developing brain: a transcriptomic, functional and ultrastructural approach // PLoS One, 2013, 8(7): 1-19, https://doi.org/10.1371/journal.pone.0065629
6. Stolp H.B., Liddelow S.A., Saunders N.R. Editorial: ontogeny and phylogeny of brain barrier mechanisms // Frontiers in Neuroscience, 2016, 10: 41, 1-3, doi: 10.3389/fnins.2016.00041.
7. Batarfi M., Valasek P., Krejci E., Huang R., Patel K. The development and origins of vertebrate meninges // Biological Communications, 2017, 62(2): 73-81, doi:10.21638/11701/spbu03.2017.203.
8. Vandenabeele F., Creemers J., Lambrichts I. Ultrastructure of the human spinal arachnoid mater and dura mater // Journal of Anatomy, 1996, 189: 417-430, PMID: 8886963.
9. Nabeshima S., Reese T., Landis D., Brightman M.W. Junctions in the meninges and marginal glia // Journal of Comparative Neurology, 1975, 164(2): 127-169, https://doi.org/10.1002/cne.901640202
10. Dias M., Mapunda J., Vladymyrov M. Structure and junctional complexes of endothelial, epithelial and glial brain barriers // International Journal of Molecular Science, 2019, 20(21): 1-27, https://doi.org/10.3390/ijms20215372
11. Pietilä R., Del Gaudio F., He L., Vázquez-Liébanas E.Vanlandewijck M., Muhl L., Mocci G., Bjørnholm K.D., Lindblad C., Fletcher-Sandersjöö A., Svensson M., Thelin E.P., Liu J. Molecular anatomy of adult mouse leptomeninges // Neuron, 2023. 111(23): 3745-3764, doi: 10.1016/j.neuron.2023.09.002
12. Wolff N. Studies on the ultrastructure of the encephalic meninges of the domestic fowl (Gallus gallus domesticus) // Digital Dissertation, 2002, 115 p.
13. Hüseynova Ş.Ə., Sadiqi İ.B., Quliyeva N.T., Yıldırım L.E., Qasımov E.K., Bağırova S.A. Ev qazlarında baş beynin sərt qişasının təşkilində iştirak edən hüceyrəvi və fibrilyar strukturların mikroskopik və ultrastruktur quruluş xüsusiyyətləri // Azərbaycan Tibb Jurnalı, 2024, №1: 142-149, https://doi.org/10.34921/amj.2024.1.022
14. Polak-Kraśna K., Robak-Nawrocka S., Szotek S. The denticulate ligament - Tensile characterisation and finite element micro-scale model of the structure stabilising spinal cord // Journal of the Medicinal Behaviour of Biomedical Materials, 2019, 91: 10-17, https://doi.org/10.1016/j.jmbbm.2018.11.017
15. Fana M., Deamont C., Medani K. Retro-Odontoid intradural synovial cyst decompression via endoscopic-assisted far-lateral approach C1-C2 hemilaminectomy without fusion: the use of intracranial denticulate ligament as intraoperative landmark // The cureus journal of medical science, 2022, 29, 14(1): 21715, doi: 10.7759/cureus.21715.
16. Nicholas D.S., Weller R.O. The fine anatomy of the human spinal meninges. A light and scanning electron microscopy study // Journal Neurosurg, 1988, 69(2): 276-282, https://doi.org/10.3171/jns.1988.69.2.0276
17. Ceylan D., Tatarlı N., Abdullaev T. The denticulate ligament: anatomical properties, functional and clinical significance // Acta Neurochirurgica, 2012, 154(7): 1229-1234, https://doi.org/10.1007/s00701-012-1361-x
18. Seragioli R., Simao M.N., Simao G.N. Assessment of the cervical spine denticulate ligament using MRI volumetric sequence: Comparison between 1.5 Tesla and 3.0 Tesla // Journal of Neuroradiology, 2018. 45(2): 147-151, https://doi.org/10.1016/j.neurad.2017.06.009
19. Elvan Ö., Kayan G., Aktekin M. The anatomical features of denticulate ligament in human fetuses // Surgical and Radiologic Anatomy, 2020, 42(8): 969-973, https://doi.org/10.1007/s00276-020-02456-0
20. Decimo I., Fumagalli G., Berton V., Krampera M., Bifari F. Meninges: rom protective membrane to stem cell niche // American Journal of Stem Cells, 2012, 1(2): 92-105, PMID: 23671802.
21. Bifari F., Berton V., Pino A., Kusalo M., GiorgioM., Di Chio M., Bersan E., Amato E., Scarpa A., Krampera M. Meninges harbor cells expressing neural precursor markers during development and adulthood // Frontiers in Cellular Neuroscience, 2015, 9, 383: 1-11, https://doi.org/10.3389/fncel.2015.00383
22. Ghannam J.Y., Al Kharazi, K.A. Neuroanatomy, Cranial Meninges // Treasure Island, Stat Pearls, 2024, 48 p, PMID: 30969704.
23. Haines D.E., Harkey H.L., Al-Mefty О. The “subdural” space: a new look at an outdated concept // Neurosurgery, 1993, 32(1): 111-120.
24. Rascher G., Wolburg H. The tight junctions of the leptomeningeal blood-cerebrospinal fluid barrier during development // The Journal fur Hirnforschung, 1997, 38(4): 525-540, PMID: 9476217.
25. Spera I., Cousin N., Ries M., Kedracka A., Castillo A., Aleandri S., Vladymyrov M., Mapunda J.A., Engelhardt B., Luciani P. Open pathways for cerebrospinal fluid outflow at the cribriform plate along the olfactory nerves // EBio Medicine, 2023, 91, 104558: 1-18, https://doi.org/10.1016/j.ebiom.2023.104558
26. Huseynova Sh., Alyarbayova A., Ghaffarzadagan R., Gasimov E. Structural features of denticulate ligaments spinal cord. Implications for transport cerebrospinal fluid // Journal of the Neurological Sciences, 2013, 333: 727.
27. Proulx S.T. Cerebrospinal fluid outflow: a review of the historical and contemporary evidence for arachnoid villi, perineural routes, and dural lymphatics // Cellular and Molecular Life Sciences, 2021, 78(6): 2429-2457, https://doi.org/10.1007/s00018-020-03706-5
28. Гасымов Э.К., Оруджев М.Т. Взаимоотношение структурных элементов оболочек, нервных волокон и кровеносных сосудов в области субарахноидального угла зрительного нерва белой крысы // Медицинский Журнал "VITA", 2001, №3-4: 16-22.
29. Барон М., Майорова Н. Функциональная стерео-морфология мозговых оболочек: Атлас // Медицина, 1982, 350 с.
30. Ichikawa H., Itoh K. Blood-arachnoid barrier disruption in experimental rat meningitis detected using gadolinium-enhancement ratio imaging // Brain Research, 2011, 16(1390): 142-149, https://doi.org/10.1016/j.brainres.2011.03.035
31. Zhang Z., Tachikawa M., Uchida Y., Terasaki T. Drug Clearance from cerebrospinal fluid mediated by organic anion transporters 1 (Slc22a6) and 3 (Slc22a8) at arachnoid membrane of rats // Molecular Pharmacology, 2018, 15(3): 911-922, https://doi.org/10.1021/acs.molpharmaceut.7b00852
32. Takeuchi H., Suzuki M., Goto R., Tezuka K., Fuchs H., Ishiguro N., Terasaki T., Braun C., Uchida Y. Regional Differences in the absolute abundance of transporters, receptors and tight junction molecules at the blood-arachnoid barrier and blood-spinal cord barrier among cervical, thoracic and lumbar spines in dogs // Pharmaceutical Research, 2022, 39(7): 1393-1413, https://doi.org/10.1007/s11095-022-03275-1
33. Verkhratsky A., Pivoriūnas A. Astroglia support, regulate and reinforce brain barriers // Neurobiology of Disease, 2023, 179: 1-11, https://doi.org/10.1016/j.nbd.2023.106054
34. Гасымов Э.K., Гусейнова Ш.А., Рзаев Ф.Г. Ультраструктурные особенности формирования структур выполняющих барьерную функцию в составе оболочек головного мозга у белых крыс // Журнал Морфология, 2020, Том. 157, №2-3: 56.
35. Smyth L.C.D., Xu D., Okar S.V., Dykstra T., Rustenhoven J., Papadopoulos Z., Bhasiin K., Kim M.W., Drieu A., Mamuladze T. Identification of direct connections between the dura and the brain // Nature, 2024, 627(8002): 165-173, https://doi.org/10.1038/s41586-023-06993-7
36. Sangha V., Aboulhassane S., Bendayan R. Regulation of folate transport at the mouse arachnoid barrier // Fluids Barriers CNS, 2024, 27, 21(1): 67, https://doi.org/10.1186/s12987-024-00566-0
Article Info:
Publication history
Published: 30.Apr.2025
Copyright
© 2022-2025. Azerbaijan Medical University. E-Journal is published by "Uptodate in Medicine" health sciences publishing. All rights reserved.Related Articles
Viewed: 2490